刘雪妮,王 超,程艳红,杨 鹏
慢性髓系白血病(chronic myeloid leukemia,CML)是早期多能造血干细胞来源的恶性骨髓增生性疾病,它的细胞遗传学的特点是存在费城(Ph)染色体t(9;22)(q34;qll)的相互易位,这些染色体的易位诱导形成了一个独特的BCR-ABL基因,随后ABL蛋白的失调导致CML细胞增殖能力增强,抗凋亡能力减弱,并改变关键的黏附性。CML在癌症治疗历史上的一个重大进展就是甲磺酸伊马替尼(imatinib mesylate, IM)的出现,它是一种半特异性的酪氨酸激酶抑制剂(tyrosine-kinase inhibitor,TKI),可以有效减少BCR-ABL mRNA的表达,并产生极高的完全血清学反应(CHR)和细胞遗传学反应(CCyR)。但是类似于其他化疗药物,IM也会出现耐药性,已知的耐药机制主要有BCR-ABL过表达与BCR-ABL扩增相关,以及耐多药P-糖蛋白(MDR-1)过表达,更常见的机制是在BCR-ABL激酶结构域的点突变的存在。GADD45α基因是一种重要的抑癌基因,在DNA修复、细胞周期阻滞、凋亡、血管生成抑制、衰老、信号转导和DNA去甲基化中发挥重要作用,GADD45α缺失可加速BCR-ABL驱动的CML,导致更具侵袭性的疾病发展。该课题组前期研究表明,GADD45α基因上游起负调控作用的miR-362-5p在IM耐药的K562细胞(K562IR)中的表达是在IM敏感的K562细胞中的4.2倍。因此,该研究推测CML细胞K562对IM的耐药可能与GADD45α表达调控有关,该研究通过过表达基因等方法,探讨GADD45α对耐药株K562IR细胞的影响。
1
.1 试剂与仪器
人慢性髓系白血病细胞K562、 IM耐药细胞K562IR购自中国科学院上海细胞生物学研究所,胎牛血清(fetal bovine serum,FBS)及RPMI 1640培养液均购自美国Gibco公司,青霉素-链霉素(PS)购自美国Sigma公司,CCK-8试剂盒购自日本Dojindo Molecular Technologies公司,慢病毒载体(滴度9×10TU/ml)及对照载体(滴度2×10TU/ml)均购自上海Genepharma公司,嘌呤霉素购自北京索莱宝科技有限公司,逆转录试剂盒购自美国Invitrogen公司,TRIzol试剂购自美国Invitrogen公司,Annexin V、PI细胞凋亡抗体购自美国BD公司,RIP裂解液、蛋白酶抑制剂及BCA蛋白定量检测试剂盒均购自上海碧云天生物技术公司,Anti-GADD45α兔单克隆抗体和HRP标记的二抗购自德国Santa Cruz Biotechnology公司,荧光显微镜购自日本Olympus公司,Western blot电泳仪和凝胶成像分析系统购自美国Bio-Rad公司,流式细胞仪、流式细胞分析软件和实时荧光定量PCR仪(qRT-PCR)购自美国BD公司。1
.2 细胞培养及转染
细胞复苏后培养于1640完全培养基(10% FBS,1% PS和RPMI 1640的培养液),37 ℃,5% CO饱和湿度下培养,适时换液、传代。取对数期的细胞进行转染。将K562和K562IR细胞按4×10/孔接种于24孔板内。设置K562不做转染作为对照组、K562IR分别转染慢病毒空载和含GADD45α的慢病毒作为阴性对照组和实验组,分别命名为K562、Lv-NC和Lv-GADD45α,将慢病毒稀释在细胞维持培养基中加入聚凝胺(5 μg/ml),后加入到相应的细胞孔板中,12~24 h移去细胞侵染后的病毒液,加入完全培养基,37 ℃,5% CO过夜,72 h后荧光显微镜观察GFP表达情况。1
.3 稳定表达细胞系的筛选
为了筛选出稳定表达的细胞株,将慢病毒载体中设计Puromycin抗性,将细胞置于含有2 μg/ml的嘌呤霉素的新鲜培养基中,每孔细胞密度不超过25%,每隔2~3 d移除和更换含有嘌呤霉素的培养基,并进行细胞观察,7 d后再次观察GFP表达情况,并收集细胞进行下一步实验。1.4 GADD45α表达的检测
① qRT-PCR检测GADD45α的mRNA水平:收集处于对数生长期的各组细胞,PBS洗2~3次后加入TRIzol试剂1 ml裂解15 min,加入三氯甲烷1 ml,低温下12 000 r/min 离心15 min,加入等体积的异丙醇混合后,静置10 min,低温下12 000 r/min离心15 min后取沉淀,紫外分光光度计下检测RNA纯度。合成引物由上海生工生物工程公司提供,序列引用先前实验,GADD45α Forward:5′-GGGCGAATTCGGATC CGCCACCATGACTTTGGAGG-3′,Reverse:5′-TTGGA ATTCGCGGCCGCTCACCGTTCAGGGAGATTAAT-3′;GAPDH Forward:5′-CGGAGTCAACGGATTTGGTCG TAT-3′,Reverse:5′-AGCCTTCTCCAT-GGTGGTGAAG AC-3′。反应条件为:95 ℃预变性5 min,95 ℃、10 s,60 ℃、30 s,72 ℃、15 s,循环40次。② Western blot检测细胞中GADD45α蛋白的表达:收集细胞后加入裂解液冰上裂解30 min,3 000 r/min离心30 min后收集上清液,并使用BCA蛋白定量试剂盒进行蛋白定量,调整各组蛋白浓度后进行上样,经过SDS-PAGE凝胶电泳分离蛋白、PVDF转膜、5%脱脂牛奶封闭2 h后,加入GADD45α(1 ∶1 000)和β-actin(1 ∶10 000)一抗4 ℃孵育过夜,TBST洗膜处理3次后再加入辣根过氧化物酶HRP标记的二抗(1 ∶1 000)室温孵育2 h,TBST再次洗膜处理3次,最后用ECL发光液中激发荧光,显影并定影。分别计算出各组蛋白的相对表达量。
1.5 转染后细胞增殖能力的CCK-8法检测
采用CCK-8法检测药物对于各组细胞的半数抑制浓度IC。培养处于对数生长期的各组细胞,稀释密度为2.0×10/ml,按100 μl/孔接种于96孔板,每组设置3个复孔,设置梯度浓度的IM干预,终浓度为0.0、1.0、5.0、10.0、20.0、50.0、100.0 μmol/L,培养24 h后,加入CCK-8试剂100 μl后继续培养2 h,经过酶标仪检测450 nm处的吸光度A。并计算IC。1.6 转染后细胞凋亡的Annexin V-FITC/PI双染流式细胞术检测
加入10 μmol / L的IM处理K562和慢病毒载体处理后的K562IR,24 h收集细胞后加入Binding Buffer重悬。而后加入Annexin V-FITC混匀避光反应10 min,再加入PI染色避光反应10 min,使用流式细胞仪检测细胞凋亡。1.7 转染后细胞迁移能力的Transwell法检测
取对数期的细胞加入Transwell小室,24孔板下室加入完全培养基和10 μmol/ L IM干预,待培养24 h后取出小室,弃去培养基,甲醇固定后用0.1%的结晶紫染色,棉签擦去上层未迁移的细胞后镜下观察。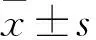
2.1 GADD45α在K562和K562IR细胞中的表达情况
采用qRT-PCR和Western blot检测K562和K562IR细胞中GADD45α的表达,结果显示K562组细胞中的GADD45α蛋白和mRNA表达水平高于K562IR细胞(P
<0.05),结果见图1。
图1 qRT-PCR和Western blot法检测K562和
2.2 转染效率检测
荧光图片显示(图2)细胞荧光效率达85%以上。采用Western blot检测转染后实验各组中GADD45α的表达,结果显示Lv-GADD45α组细胞GADD45α蛋白表达水平上调,相比Lv-NC组差异有统计学意义(P
<0.05) ,而Lv-GADD45α组和K562组比较差异无统计学意义,结果见图3。
图2 荧光显微镜下观察Lv-NC组、Lv-GADD45α组细胞 ×200

图3 Western blot检测K562组、Lv-NC组、Lv-GADD45α组中GADD45α蛋白的表达
2.3 GADD45α基因上调后细胞增殖能力的变化
利用终浓度为0.0、 1.0、5.0、10.0、20.0、50.0、100.0 μmol/L的IM干预K562、Lv-NC、Lv-GADD45α组后细胞存活率下降。3组的IC分别为10.08 μmol / L、50.56 μmol/L、18.72 μmol/L。从结果来看,Lv-GADD45α组的存活率低于Lv-NC组,结果见图4。
图4 CCK-8法检测不同浓度IM处理的各组细胞存活率
2.4 GADD45α基因上调后细胞体外凋亡的变化
以10 μmol/L的IM为测试剂量,分别干预K562、Lv-NC、Lv-GADD45α组细胞,检测在耐药剂量下细胞凋亡的试验结果。结果显示,K562组和Lv-GADD45α组凋亡率与Lv-NC组相比差异有统计学意义(P
<0.01,F
=1.44、0.96,t
=11.42、7.76),结果见图5。
图5 流式细胞仪检测K562组、Lv-NC组、Lv-GADD45α组细胞凋亡率
2.5 GADD45α基因上调后迁移能力的变化
使用等剂量10.0 μmol/L的IM对各组细胞进行干预,通过Transwell迁移小室检测各组细胞的迁移能力,观察小室底膜上下室侧附着的细胞,取6个视野进行观察和拍照,同时进行细胞计数。结果显示K562组和Lv-GADD45α组细胞穿过的数目低于Lv-NC组细胞,K562组和Lv-GADD45α组相对于Lv-NC细胞迁移能力下降(P
<0.01,F
=2.46、2.57,t
=11.89、9.01),结果见图6。
图6 Transwell法检测K562组、Lv-NC组、Lv-GADD45α组的侵袭能力 结晶紫染色×100
IM目前被认为是慢性阶段CML患者的一线治疗药物,也是CML晚期快速缓解的首选药物,尽管通常是短期缓解,然而,约有1/3的患者在接受IM治疗时会产生对IM的耐药性。而在后来出现的第二代TKl尼洛替尼(nilotinib)、达沙替尼(dasision)等虽然在治疗过程中会比IM出现更低的初级耐药和突变发生,但尚未转化为长期的总体生存优势。越来越多的证据表明,并非所有的IM耐药都来自BCR-ABL激酶结构域突变,已有研究表明,血红素加氧酶-1通过干扰哺乳动物雷帕霉素靶蛋白(mTOR)信号通路诱导自噬形成,导致CML发生IM耐药。因此,研究这些非突变机制在CML耐药中具有重要意义。
GADD45α是一种广泛表达的核蛋白,与参与DNA修复、细胞周期调节、凋亡、衰老和自噬的细胞蛋白发生物理作用,并且它的缺失显示了基因组的不稳定性和增加了对致癌物的敏感性,促进肿瘤的发生。GADD45α在维持造血干细胞基因组完整性方面的作用是至关重要的,而其作为血液恶性肿瘤的肿瘤抑制因子的作用模式,除了提供DNA修复和基因组稳定性外,诱导分化则是GADD45α在造血系统中的主要抗白血病作用。GADD45α的上调是多种化疗药物抗肿瘤活性的重要步骤,例如多西紫杉醇、姜黄素等,而GADD45α的沉默可能会削弱这些药物的治疗效果。该课题组前期研究表明,miR-362-5p是GADD45α反向调控的miRNA,其在K562IR中的表达高于K562细胞中的表达,当IM抑制BCR-ABL活性后可降低K562细胞中miR-362-5p的表达水平。因此,该研究推测在慢性粒细胞白血病中对IM的耐药可能与GADD45α的表达有关。
该研究考察了K562和K562IR细胞中GADD45α的mRNA和蛋白的表达的情况,GADD45α在IM敏感的K562 细胞中高表达,高于K562IR细胞,这与之前的研究结果一致,提示GADD45α可能参与了细胞发生耐药的过程。进一步通过慢病毒转染的方法上调了耐药细胞株中 GADD45α基因,并通过Western blot检测细胞转染效率,结果显示相比对照组和Lv-NC组,Lv-GADD45α组中GADD45α的表达上调。同时为了进一步研究上调GADD45α对K562IR的影响,该研究通过不同浓度的IM对各分组细胞进行干预,检测不同组细胞的存活率、凋亡率和侵袭能力,结果表明Lv-GADD45α细胞组的凋亡率增高,存活率和侵袭能力降低。结果显示,GADD45α的低表达可能是慢性髓细胞白血病细胞K562产生耐药的一种机制。
已有研究表明,在BCR-ABL转化髓系祖细胞中,当GADD45α缺失时会高度激活p38MAPK、PI3K/AKT和STAT5信号通路,促进细胞的增殖和减少凋亡。因此,当GADD45α基因上调后,可能是通过抑制PI3K/AKT/mTOR信号通路降低了表达BCR/ABL的慢性髓系白血病细胞的耐药性,具体的机制还需进一步研究。